
16 Neurotransmitters: Amino Acid Neurotransmitters (Glutamate, GABA, Glycine)
Glutamate
Glutamate is an amino acid transmitter and is the primary excitatory neurotransmitter in the brain. Glutamate is the same as the amino acid glutamic acid. There is more glutamate per volume of brain tissue than any other neurotransmitter.
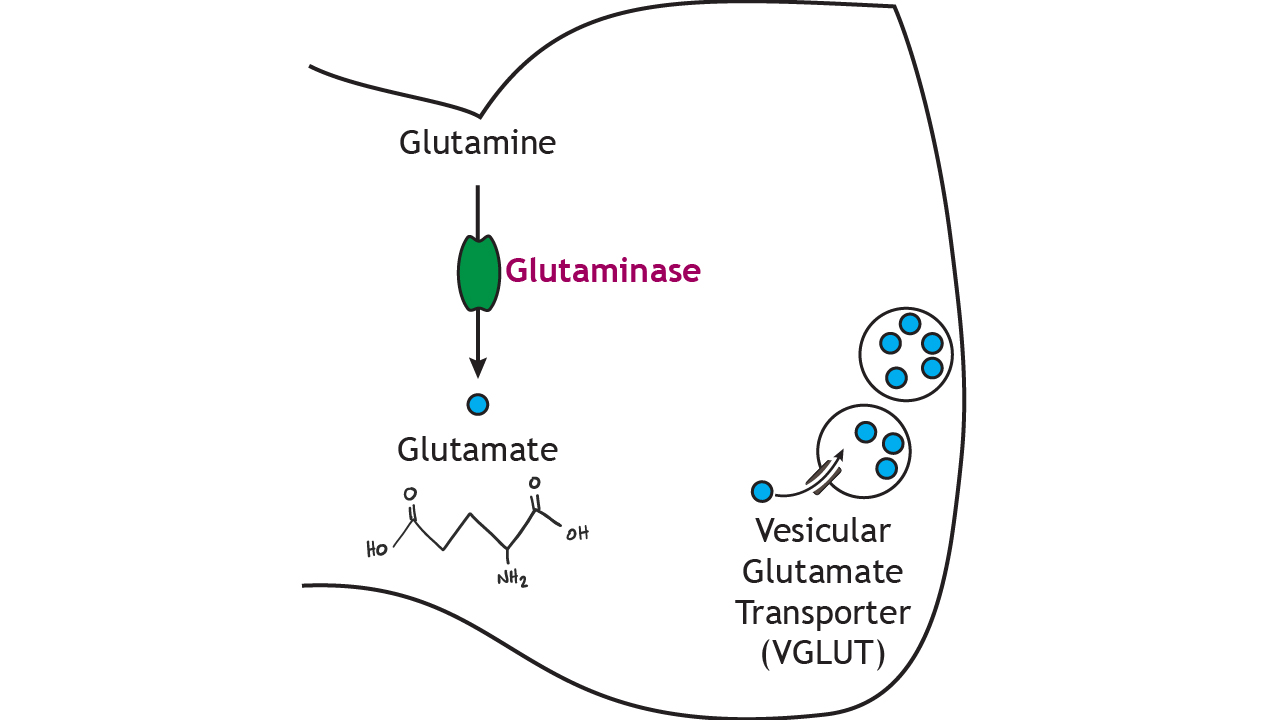
In the presynaptic terminal, glutamine is converted into glutamate via the enzyme glutaminase, which is the rate-limiting step in the synthesis pathway. Glutamate is packaged into small vesicles for storage via the vesicular glutamate transporter (vGLUT). Staining for the presence of vGLUT is one way that researchers are able to identify glutamatergic neurons.
Glutamate Receptors
Glutamate can activate both ionotropic and metabotropic receptors. Glutamate is the primary excitatory neurotransmitter in the central nervous system and opens non-selective cation channels. There are three subtypes of ionotropic glutamate receptors: AMPA, kainate, and NMDA receptors.
The AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) and kainate receptors allow both sodium and potassium to cross the membrane. Although potassium can leave the cell when the receptors open, the electrochemical gradient driving sodium ion movement is stronger than the gradient driving potassium movement, resulting in a depolarization of the membrane potential.
Animation 16.1. AMPA and kainate glutamate receptors are non-selective ion channels that allow both sodium and potassium to flow across the membrane. When glutamate binds, sodium flows in and potassium flows out. The lined, teal channel represent AMPA receptors; the checkered, teal channel represents kainate receptors. ‘AMPA and Kainate’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
The NMDA (N-methyl-D-aspartate) receptor requires the binding of glutamate to open, but it is also dependent on voltage. When the membrane potential is below, at, or near rest, a magnesium ion blocks the open NMDA receptor and prevents other ions from moving through the channel. Once the cell depolarizes, the magnesium block is expelled from the receptor, which allows sodium, potassium, and calcium to cross the membrane. The voltage change needed to open the NMDA receptor is usually a result of AMPA receptor activation.
Animation 16.2. NMDA receptors are opened by a combination of glutamate binding and a voltage trigger. At low levels of stimulation, when the the membrane potential is near rest, a magnesium ion blocks the open NMDA receptor channel preventing ion flow. Ions can flow through open AMPA receptors, which begins to depolarize the membrane. The voltage change eventually expels the magnesium ion from the channel, allowing sodium, potassium, and calcium to cross the membrane. The lined, teal channel represents AMPA receptors; the dotted, violet channel represents NMDA receptors. ‘AMPA and NMDA’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Termination of Glutamate Signaling
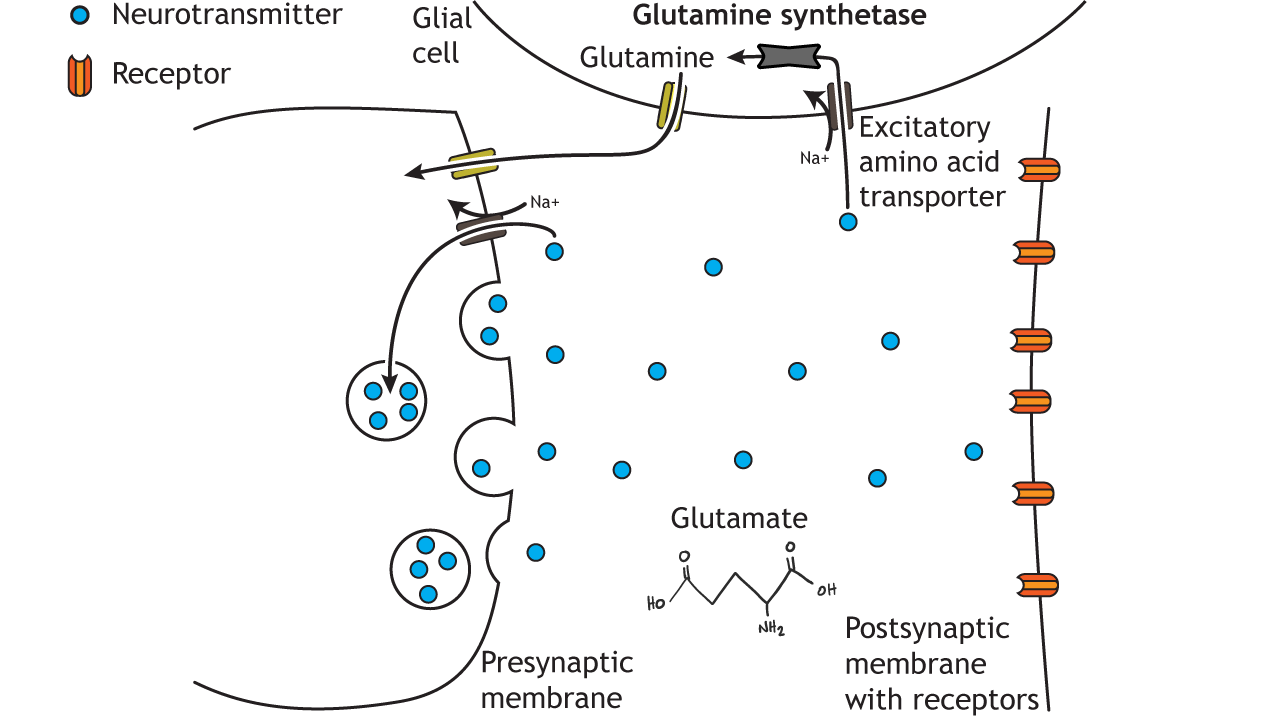
Glutamate action is terminated by two mechanisms. Reuptake of glutamate molecules into the presynaptic terminal can occur, or glutamate can be transported into nearby glial cells. The excitatory amino acid transporters are sodium co-transporters and use the sodium electrochemical gradient to drive neurotransmitter transport. Within glial cells, glutamate is converted into glutamine by glutamine synthetase. Glutamine is then transported out of the glial cell and back into the presynaptic terminal for use in future glutamate synthesis. If glutamate is transported back into the presynaptic terminal, it can be repackaged in synaptic vesicles.
GABA
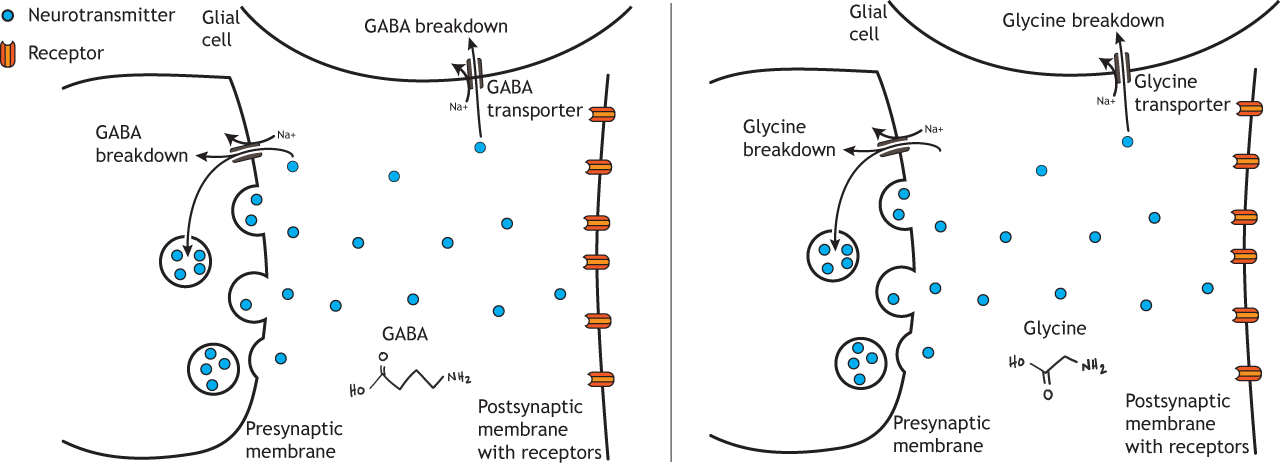
Glutamate is then used to synthesize Gamma-aminobutyric acid (GABA), the main inhibitory neurotransmitter in the brain. According to one estimate, about 25% of neurons in the brain are GABAergic.
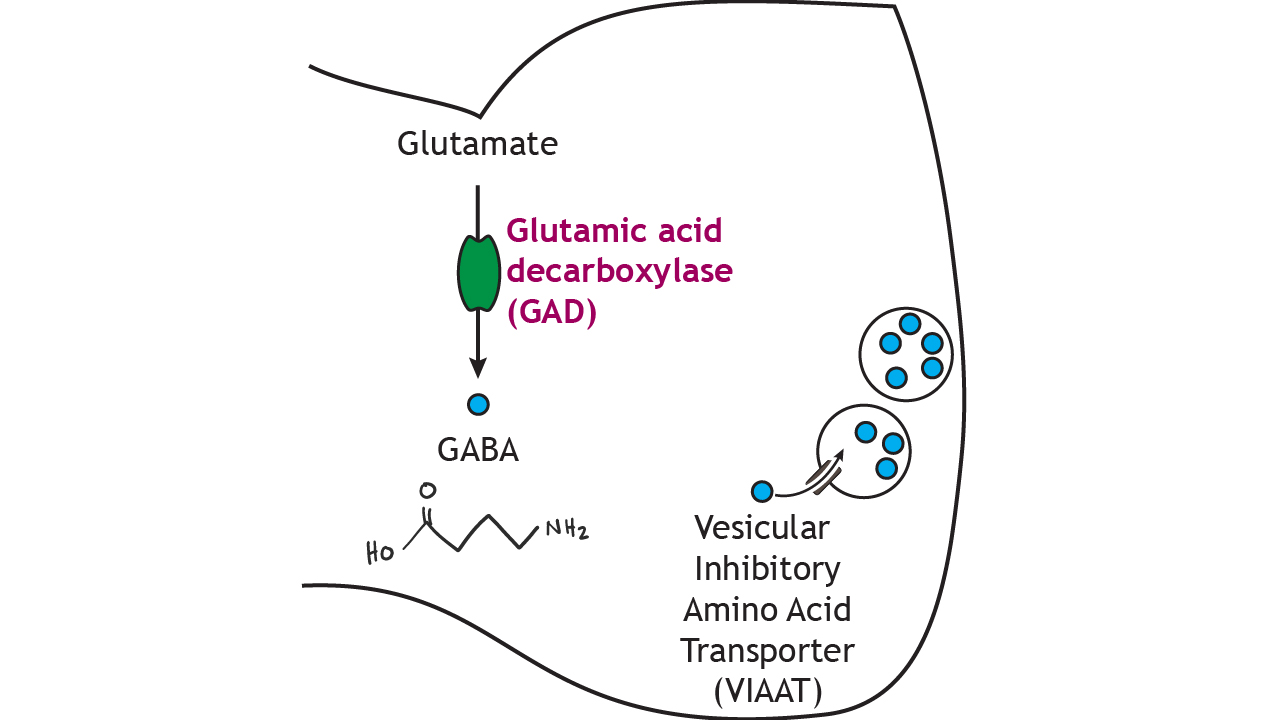
In the presynaptic terminal, glutamate is converted into GABA via the enzyme glutamic acid decarboxylase (GAD), which—like the other synthesis pathways—is the rate-limiting step. GAD is often used as a biochemical marker for the presence of GABAergic neurons. GABA is packaged into small vesicles for storage in the terminal via the vesicular inhibitory amino acid transporter (VIAAT).
Glycine
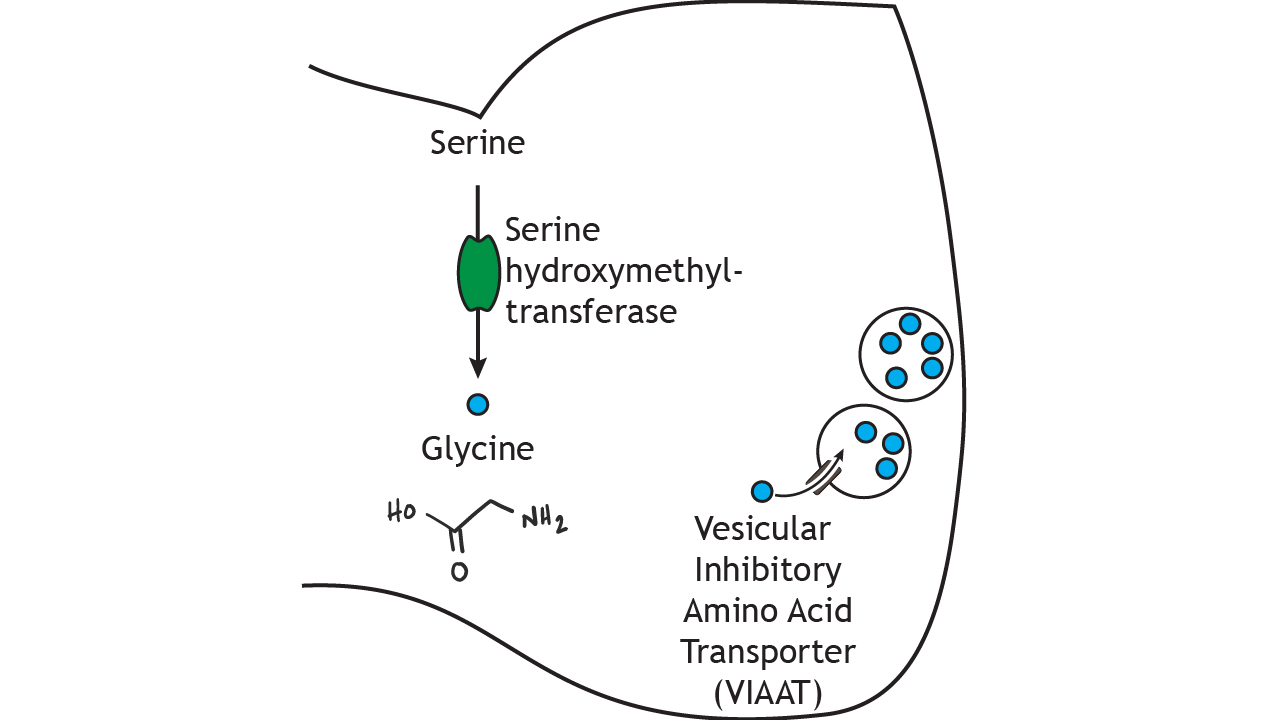
Glycine is another inhibitory amino acid neurotransmitter, but unlike GABA, is mostly used by neurons of the spinal cord and brain stem. Serine hydroxymethyltransferase converts the amino acid serine into glycine in the presynaptic terminal. The rate-limiting step for glycine synthesis occurs earlier in the pathway prior to serine synthesis. Glycine is packaged into small vesicles by the vesicular inhibitory amino acid transporter (VIAAT) like GABA.
GABA and Glycine Receptors
GABA and glycine receptors are chloride channels. Since an increase in chloride permeability across the membrane is inhibitory, the binding of GABA or glycine to their respective ionotropic receptor will cause inhibition.
Animation 16.3. GABA and glycine are inhibitory receptors that are selective to chloride. The solid yellow channel represents a GABA receptor; the patterned, yellow channel represents a glycine receptor. ‘GABA and Glycine’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC-BY-NC-SA) 4.0 International License. View static image of animation.
Termination of GABA and Glycine Signaling
Like glutamate, GABA and glycine action are terminated by either reuptake into the presynaptic terminal and packaging in synaptic vesicles or through transport into glial cells where breakdown can occur. The GABA and glycine transporter also use the sodium electrochemical gradient to drive the movement of the transmitter across the membrane.
Key Takeaways
- Glutamate is the primary excitatory neurotransmitter in the brain
- Glutamate binds to both ionotropic (AMPA, NMDA, and kainate) receptors and metabotropic receptors
- GABA is the primary inhibitory neurotransmitter in the brain
- Glycine is the primary inhibitory neurotransmitter in the periphery
- GABA and Glycine bind to ionotropic receptors that open chloride channels
Test Yourself!
Attributions
Portions of this chapter were remixed and revised from the following sources:
- Foundations of Neuroscience by Casey Henley. The original work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
primary excitatory neurotransmitter
When the membrane potential gets closer to zero
Process where neurotransmitters are removed from the synapse and transported back into the presynaptic cell to be re-packaged into synaptic vesicles
primary inhibitory neurotransmitter in the brain
