
29 Visual System: The Eye
Humans are remarkably dependent on the visual system to gain information about our surroundings. Consider how tentatively you walk from the light switch to your bed right after turning off the lights!
The visual system is complex and consists of several interacting anatomical structures. Here, we will describe the process of how photons of light from our surroundings become signals that the brain turns into representations of our surroundings.
Properties of Light
Visual sensation starts at the level of the eye. The eye is an organ that has evolved to capture photons, the elementary particle of light. Photons are unusual because they behave as both particles and as waves, but neuroscientists mostly focus on the wave-like properties. Because photons travel as waves, they oscillate at different frequencies. The frequency at which a photon oscillates is directly related to the color that we perceive.
The human visual system is capable of seeing light in a very narrow range of frequencies on the electromagnetic spectrum. On the short end, 400 nm wavelengths are observed as violet, while on the long end, 700 nm wavelengths are red. Ultraviolet light oscillates at a wavelength shorter than 400 nm, while infrared light oscillates at a wavelength longer than 700 nm. Neither ultraviolet nor infrared light can be detected with our eyes.
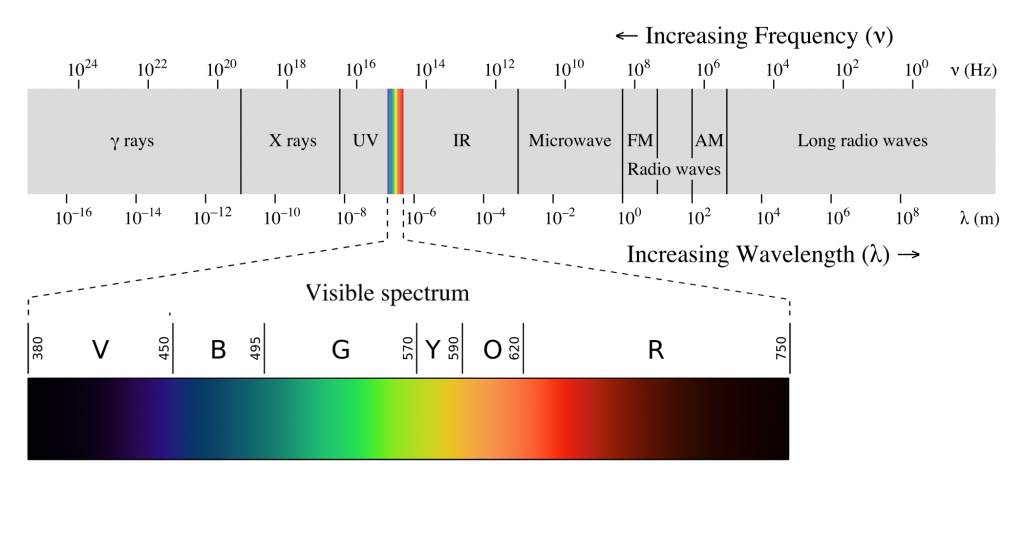
Anatomy of the Eye
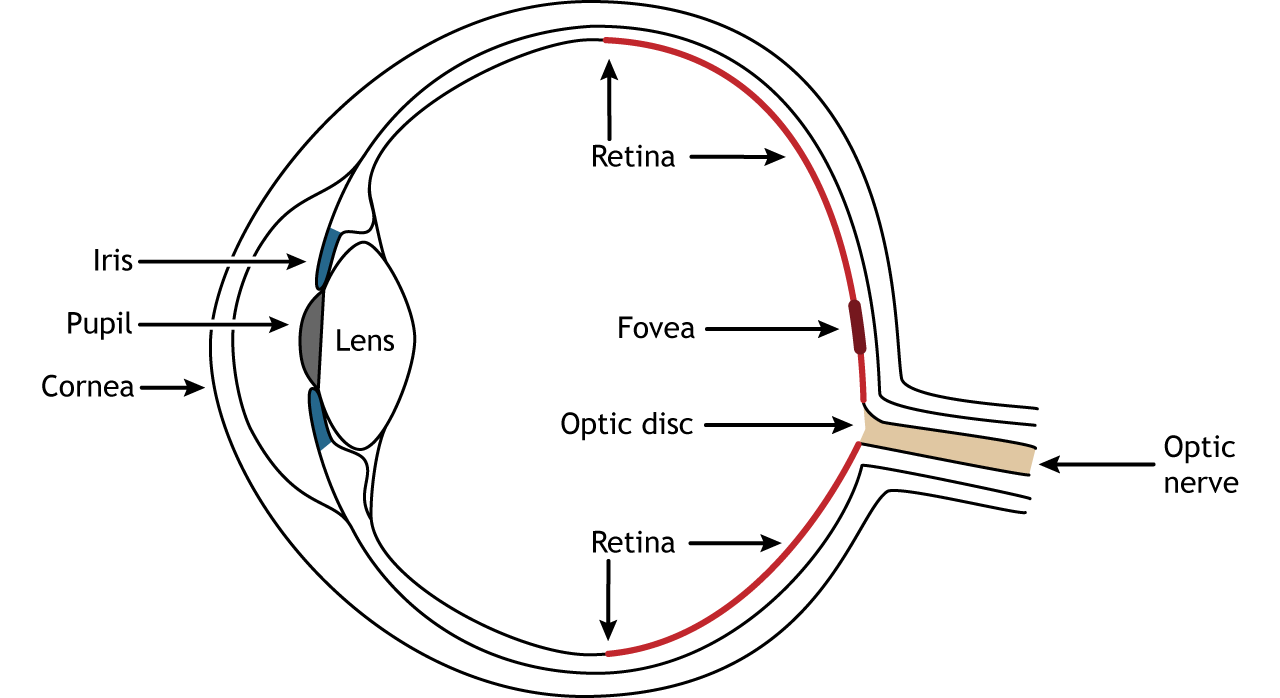
Photons pass through several anatomical structures before the nervous system processes and interprets them. The front of the eye consists of the cornea, pupil, iris, and lens. The cornea is the transparent, external part of the eye. The cornea refracts, or bends, the incoming rays of light so that they converge precisely at the retina, the posterior most part of the eye. If the light rays fail to properly converge, a person would be near-sighted or far-sighted, and this would result in blurry vision. Glasses or contact lenses bend light before it reaches the cornea to compensate the cornea’s shape.
After passing through the cornea, light enters through a hole in the opening in the iris at the center of the eye called the pupil. The iris is the colored portion of the eye that surrounds the pupil and along with local muscles that can control the size of the pupil to allow for an appropriate amount of light to enter the eye. The diameter of the pupil can change depending on ambient light conditions. In the dark, the pupil dilates, or gets bigger, which allows the eye to capture more light. In bright conditions, the pupils constricts, or gets smaller, which decreases the amount of light that enters the eye.
The next structure that light passes through is the lens. The lens is located behind the pupil and iris. Like the cornea, the lens refracts light so that the rays converge on the retina. Proper focusing requires the lens to stretch or relax, a process called accommodation. A circular muscle that surrounds the lens, called the ciliary muscle, changes the shape of the lens depending on the distance of the object of focus.
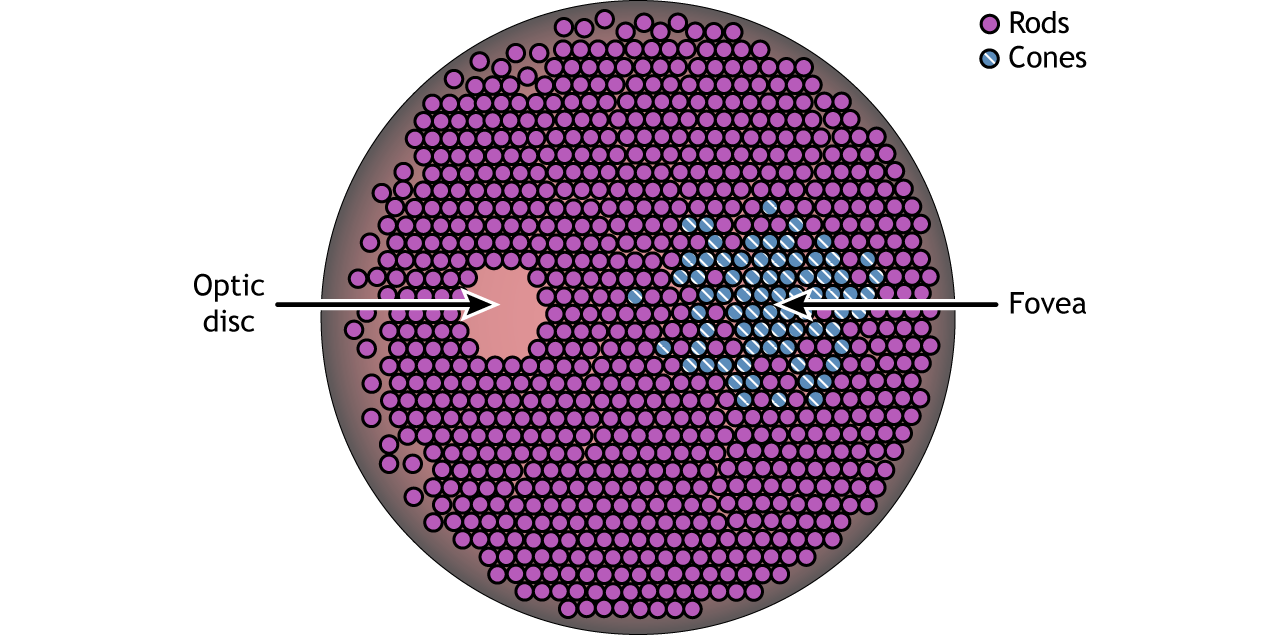
The retina is the light-sensitive region in the back of the eye where the photoreceptors, the specialized cells that respond to light, are located. The retina covers the entire back portion of the eye, so it’s shaped like a bowl. In the middle of the bowl is the fovea, the region of highest visual acuity, meaning the area that can form the sharpest images. The optic nerve projects to the brain from the back of the eye, carrying information from the retinal cells. Where the optic nerve leaves, there are no photoreceptors since the axons from the neurons are coming together. This region is called the optic disc and is the location of the blind spot in our visual field.
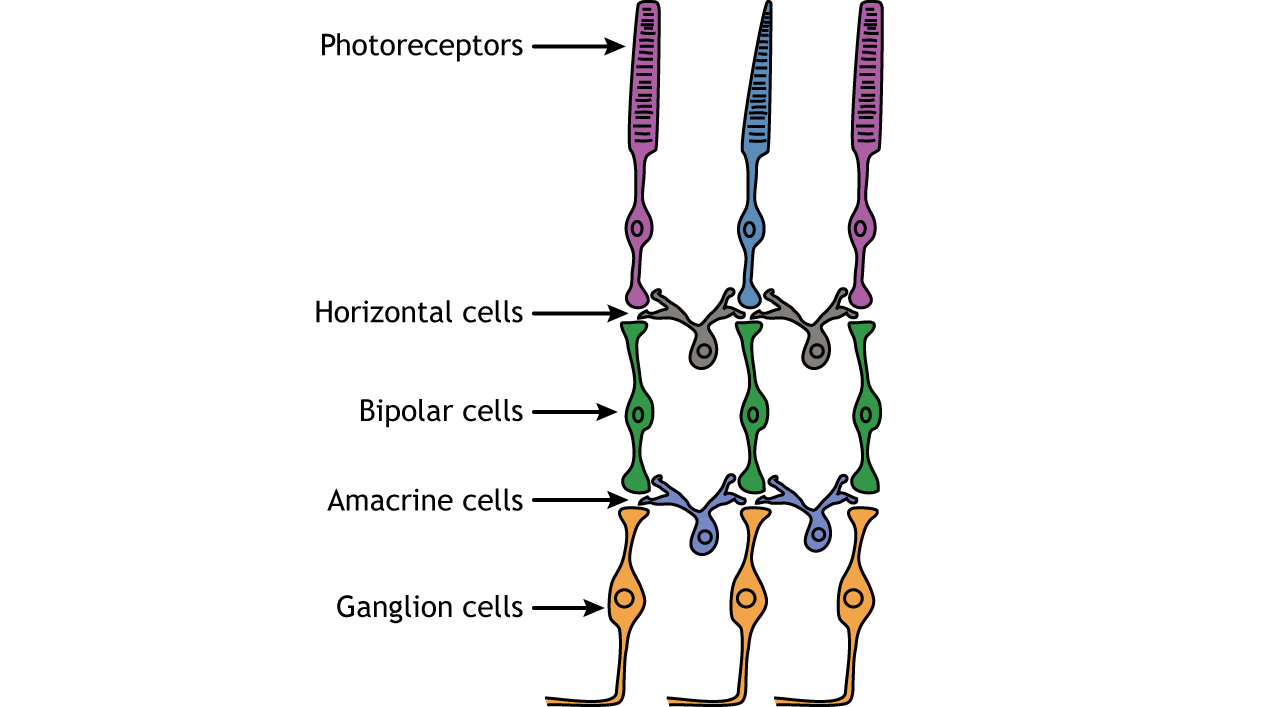
Retinal Cells
In addition to the photoreceptors, there are four other cell types in the retina. The photoreceptors synapse on bipolar cells, and the bipolar cells synapse on the ganglion cells. Horizontal and amacrine cells allow for communication laterally between the neuron layers.
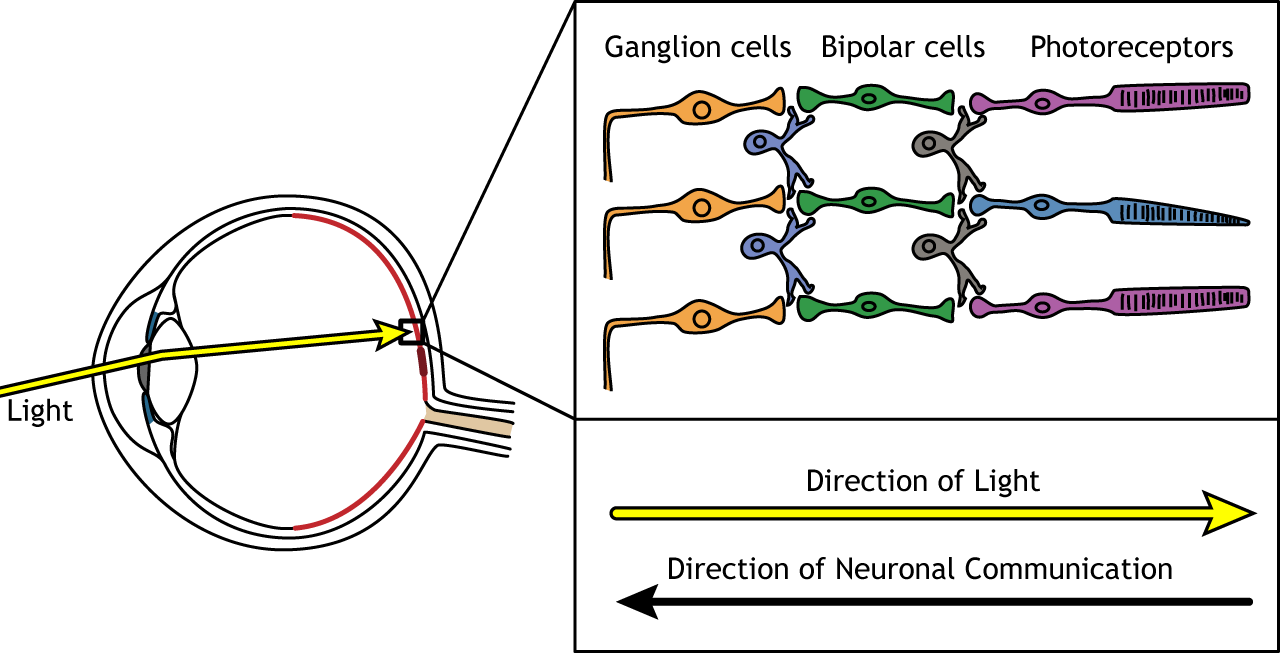
Direction of Information
When light enters the eye and strikes the retina, it must pass through all the neuronal cell layers before reaching and activating the photoreceptors. The photoreceptors then initiate the synaptic communication back toward the ganglion cells.
Photoreceptors
Photoreceptors are the first cells in the neuronal visual perception pathway. The photoreceptors are the specialized receptors that respond to light. They are the cells that detect photons of light and convert them into neurotransmitter release, a process called phototransduction.
Morphologically, photoreceptor cells have two parts, an outer segment and inner segment. The outer segment contains stacks of membranous disks bounded within the neuronal membrane. These membranous disks contain molecules called photopigments, which are the light-sensing components of the photoreceptors. Hundreds of billions of these photopigments can be found in a single photoreceptor cell. The inner segment contains the nucleus and other organelles. Extending from the inner segment is the axon terminal.
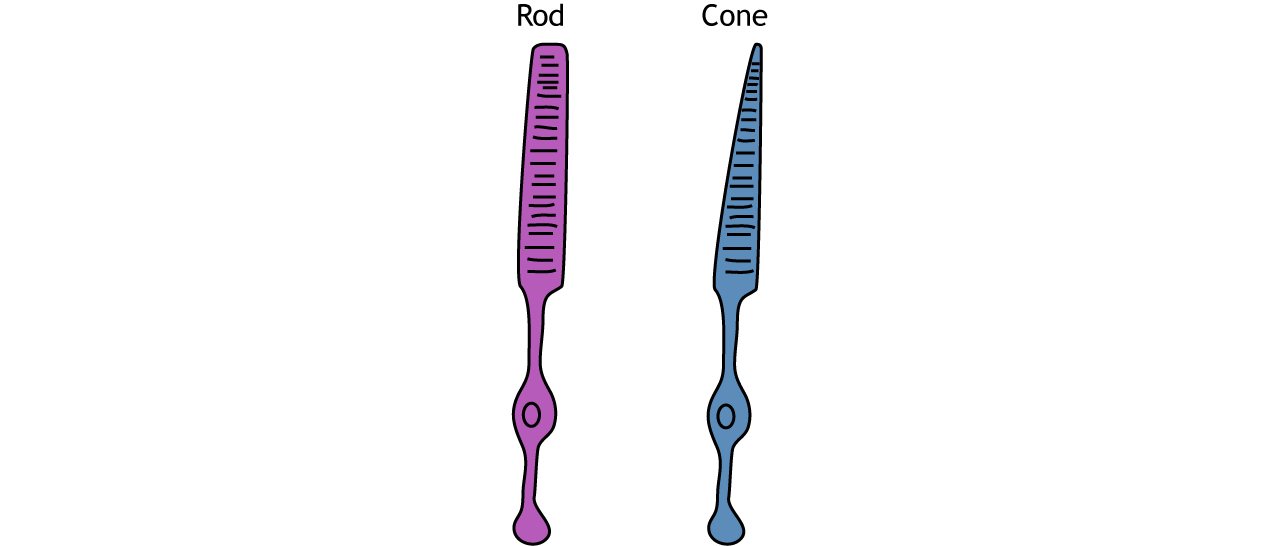
Photoreceptors are classified into two categories, named because of their appearance and shape: rods and cones. Rod photoreceptors have a long cylindrical outer segment that holds many membranous disks. The presence of more membranous disks means that rod photoreceptors contain more photopigments and thus are capable of greater light sensitivity.
Cone photoreceptors have a short, tapered c, and cylindrical outer segment that holds fewer membranous disks than rod photoreceptors. The presence of less membranous disks means that cone photoreceptors contain less photopigments and thus are not as sensitive to light as rod photoreceptors. Cone photoreceptors are responsible for processing our sensation of color (the easiest way to remember this is cones = color). The typical human has three different types of cone photoreceptors cells, with each of these three types tuned to specific wavelengths of light. The short wavelength cones (S-cones) respond most robustly to 420 nm violet light. The middle wavelength cones (M-cones) exhibit peak responding at 530 nm green light, and the long wavelength cones (L-cones) are most responsive in 560 nm red light. Each of these cones is activated by other wavelengths of light too, but to a lesser degree. Every color on the visible spectrum is represented by some combination of activity of these three cone photoreceptors.
The idea that we have two different cellular populations and circuits that are used in visual perception is called the duplicity theory of vision and is our current understanding of how the visual system perceives light. It suggests that both the rods and cones are used simultaneously and complement each other. The photopic vision, uses cone photoreceptors of the retina, and is responsible for high-acuity sight and color vision in daytime. Its counterpart, called scotopic vision, uses rod photoreceptors and is best for seeing in low-light conditions, such as at night. Both rods and cones are used for mesopic vision, when there are intermediate lighting conditions, such as indoor lighting or outdoor traffic lighting at night.
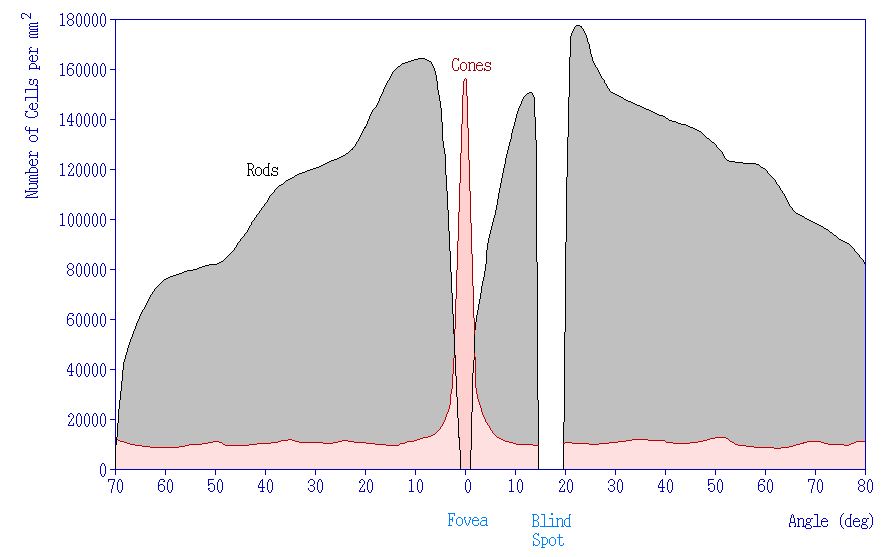
Photoreceptor Density
In addition to having different visual functions, the rods and cones are also distributed across the retina in different densities. Visual information from our peripheral vision is generally detected by our rod cells, which are most densely concentrated outside the fovea. Cone photoreceptor cells allow for high-acuity vision. They are most densely packed at the fovea, corresponding to the very center of your visual field. Despite being the cell population that we use for our best vision, cone cells make up the minority of photoreceptors in the human retina, outnumbered by about 20-times more rod cells.
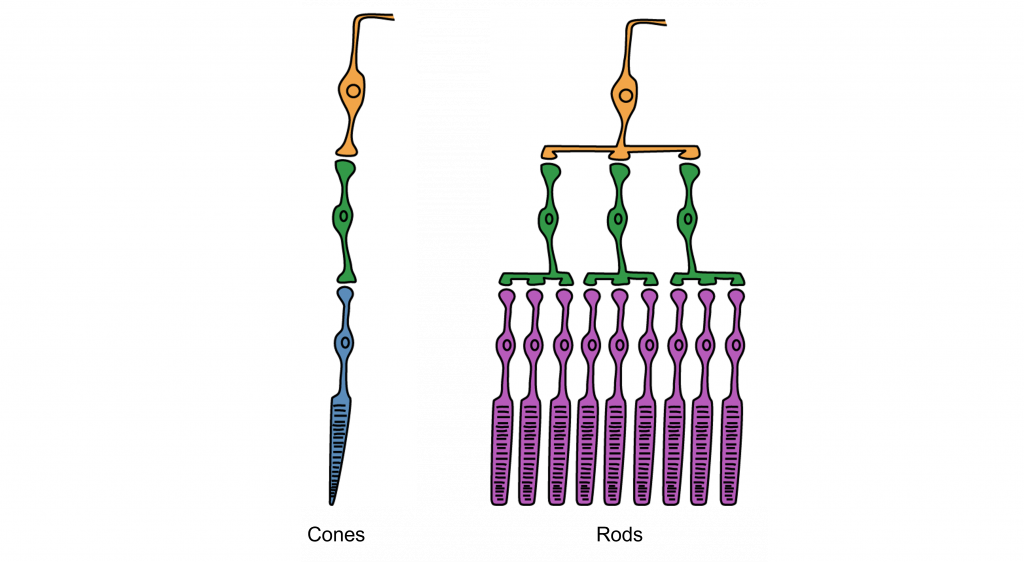
Synaptic Convergence of Photoreceptors
Rod cells are organized to have high synaptic convergence, where several rod cells (up to 30) feed into a single downstream route of communication (the bipolar cells, to be specific). An advantage of a high-convergence network is the ability to add many small signals together to create a seemingly larger signal. Consider stargazing at night, for example. Each rod is able to detect low levels of light, but signals from multiple rod cells, when summed together, allows you to recognize faint light sources such as a star. A disadvantage of this type of organization is that it is difficult to identify exactly which photoreceptor is activated by the incoming light, which is why accuracy is poor when seeing stimuli in our peripheral vision. This is one of the reasons that we cannot actually read text in our peripheral vision or see the distinct edges of a star. Rod photoreceptors are maximally active in low-light conditions.
Unlike rod cells, cone cells have very low synaptic convergence. In fact, at the point of highest visual acuity, a single cone photoreceptor communicates with a single pathway to the brain. The signaling from low-convergence networks is not additive, so they are less effective at low light conditions. However, because of this low-convergence organization, cone cells are highly effective at precisely identifying the location of incoming light.
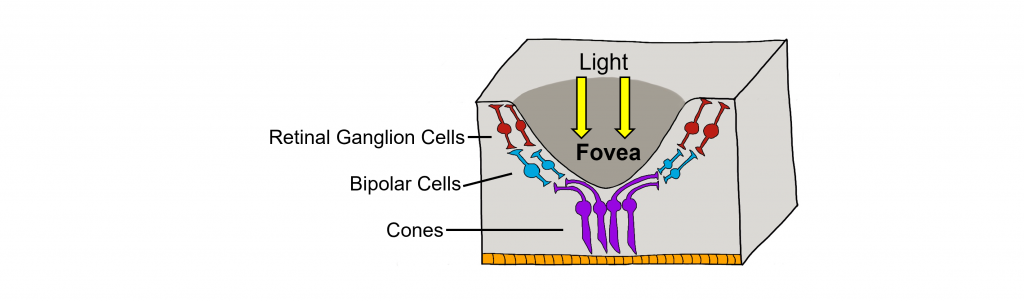
Retina: Fovea and Optic Disk
The retina is not completely uniform across the entire back of the eye. There are a few spots of particular interest along the retina where the cellular morphology is different: the fovea and the optic disk.
There are two cellular differences that explain why the fovea is the site of our best visual acuity. For one, the neurons found at the fovea are “swept” away from the center, which explains why the fovea looks like a pit. Cell membranes are made up mostly of lipids, which distort the passage of light. Because there are fewer cell bodies present here, the photons of light that reach the fovea are not refracted by the presence of other neurons. Secondly, the distribution of photoreceptors at the fovea heavily leans toward cone type photoreceptors. Because the cone cells at the fovea exhibit low convergence, they are most accurately able to pinpoint the exact location of incoming light. On the other hand, most of the photoreceptors in the periphery are rod cells. With their high-convergence circuitry, the periphery of the retina is suited for detecting small amounts of light, though location and detail information is reduced.
Another anatomically interesting area of the retina is an elliptical spot called the optic disk. This is where the optic nerve exits the eye. At this part of the retina, there is an absence of photoreceptor cells. Because of this, we are unable to perceive light that falls onto the optic disk. This spot in our vision is called the blind spot.
Phototransduction
The photoreceptors are responsible for sensory transduction in the visual system, converting light into electrical signals in the neurons. For our purposes, to examine the function of the photoreceptors, we will A) focus on black and white light (not color vision) and B) assume the cells are moving from either an area of dark to an area of light or vice versa.
Photoreceptors do not fire action potentials; they respond to light changes with graded receptor potentials (depolarization or hyperpolarization). Despite this, the photoreceptors still release glutamate onto the bipolar cells. The amount of glutamate released changes along with the membrane potential, so a hyperpolarization will lead to less glutamate being released. Photoreceptors hyperpolarize in light and depolarize in dark. In the graphs used in this lesson, the starting membrane potential will depend on the initial lighting condition.
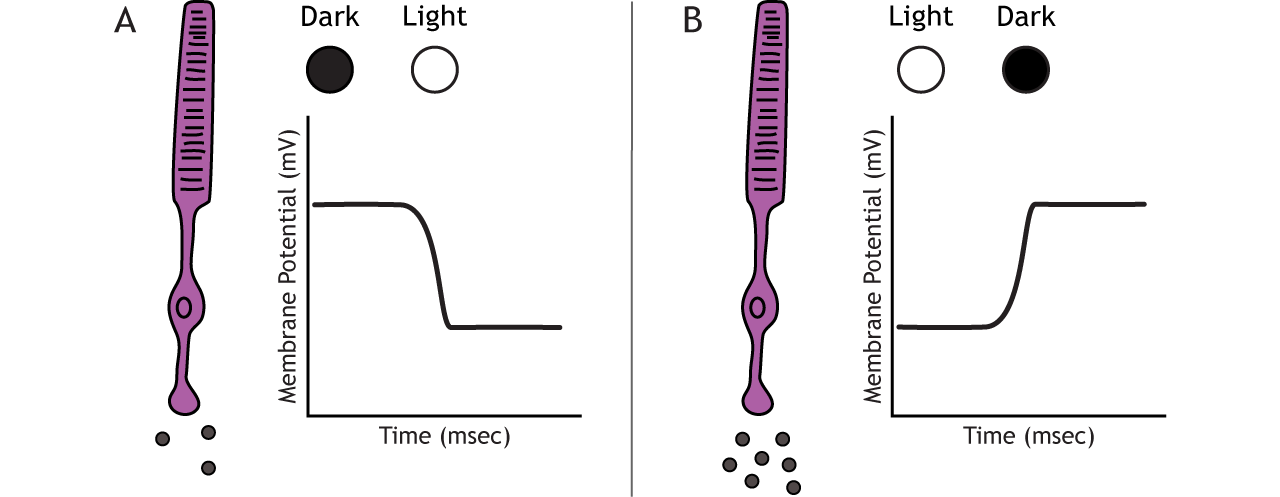
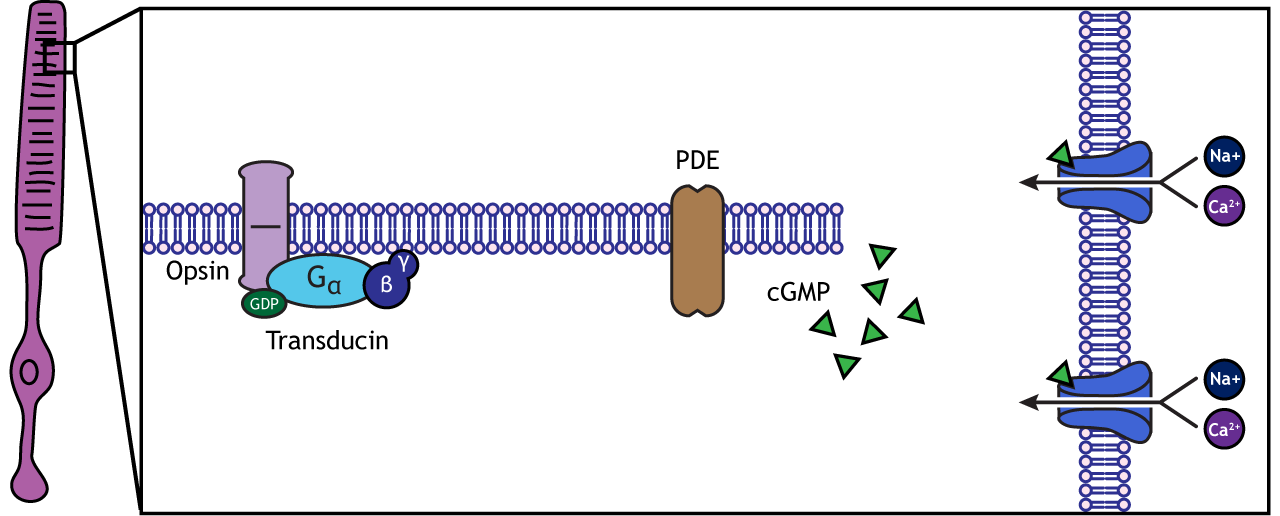
When the photoreceptor moves into the light, the cell hyperpolarizes. Light enters the eye, reaches the photoreceptors, and causes a conformational change in a special receptor protein called an opsin. The opsin receptor has a pre-bound chemical agonist called retinal. Together, the opsin + retinal makes up the photopigment rhodopsin.
When rhodopsin absorbs light, it causes a conformational change in the pre-bound retinal, in a process called “bleaching”. The bleaching of rhodopsin activates an associated G-protein called transducin, which then activates an effector enzyme called phosphodiesterase (PDE). PDE breaks down cGMP in the cell to GMP. As a result, the cGMP-gated ion channels close. The decrease in cation flow into the cell causes the photoreceptor to hyperpolarize.
Animation 29.1. Light reaching the photoreceptor causes a conformational change in the opsin protein, which activates the G-protein transducing. Transducin activates phosphodiesterase (PDE), which converts cGMP to GMP. Without cGMP, the cation channels close, stopping the influx of positive ions. This results in a hyperpolarization of the cell. ‘Phototransduction’ by Casey Henley is licensed under a Creative Commons Attribution Non-Commercial Share-Alike (CC BY-NC-SA) 4.0 International License. View static image of animation.
In the dark, the photoreceptor has a membrane potential that is more depolarized than the “typical” neuron we examined in previous chapters; the photoreceptor membrane potential is approximately -40 mV.
Rhodopsin is not bleached, thus the associated G-protein, transducin, remains inactive. As a result, there is no activation of the PDE enzyme, and levels of cGMP within the cell remain high. cGMP binds to cGMP-gated sodium ion channels, causing them to open. The open cation channels allow the influx of sodium and calcium, which depolarize the cell in the dark.
Transmission of Information within Retina
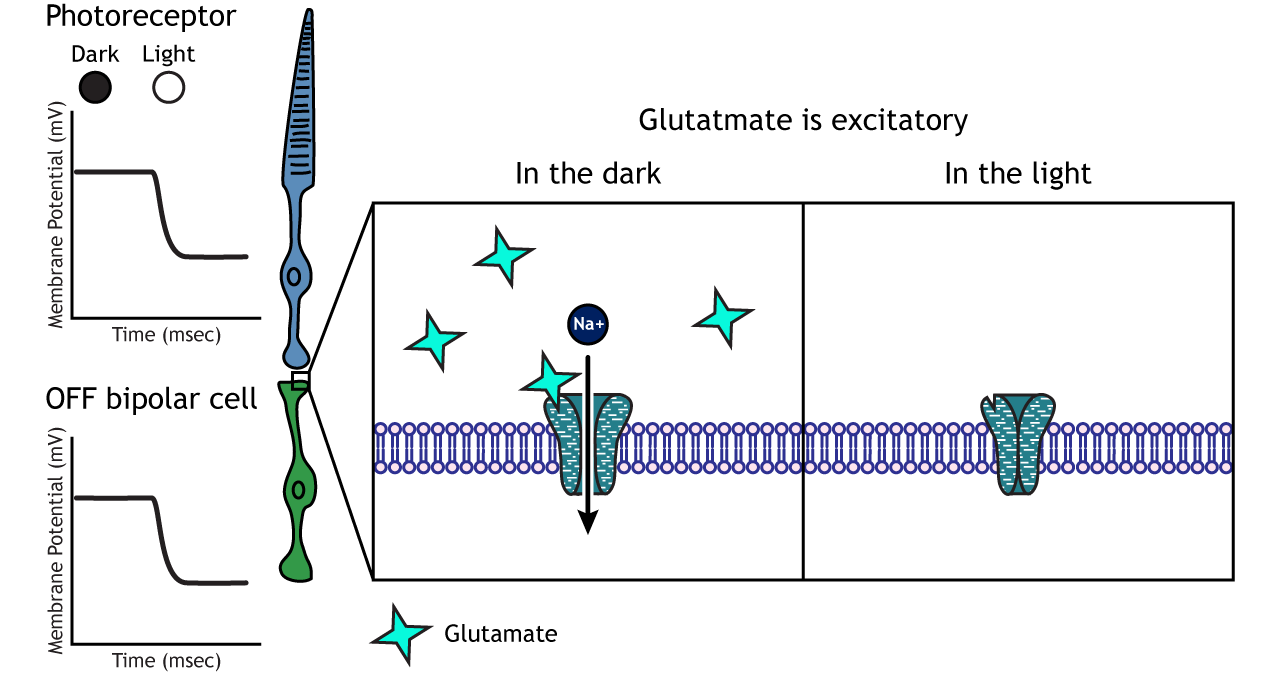
Photoreceptors synapse onto bipolar cells in the retina. There are two types of bipolar cells: OFF-center bipolar cells and ON-center bipolar cells. These cells respond in opposite ways to the glutamate released by the photoreceptors because they express different types of glutamate receptors. Like photoreceptors, the bipolar cells do not fire action potential and only respond with graded postsynaptic potentials.
OFF-center Bipolar Cells
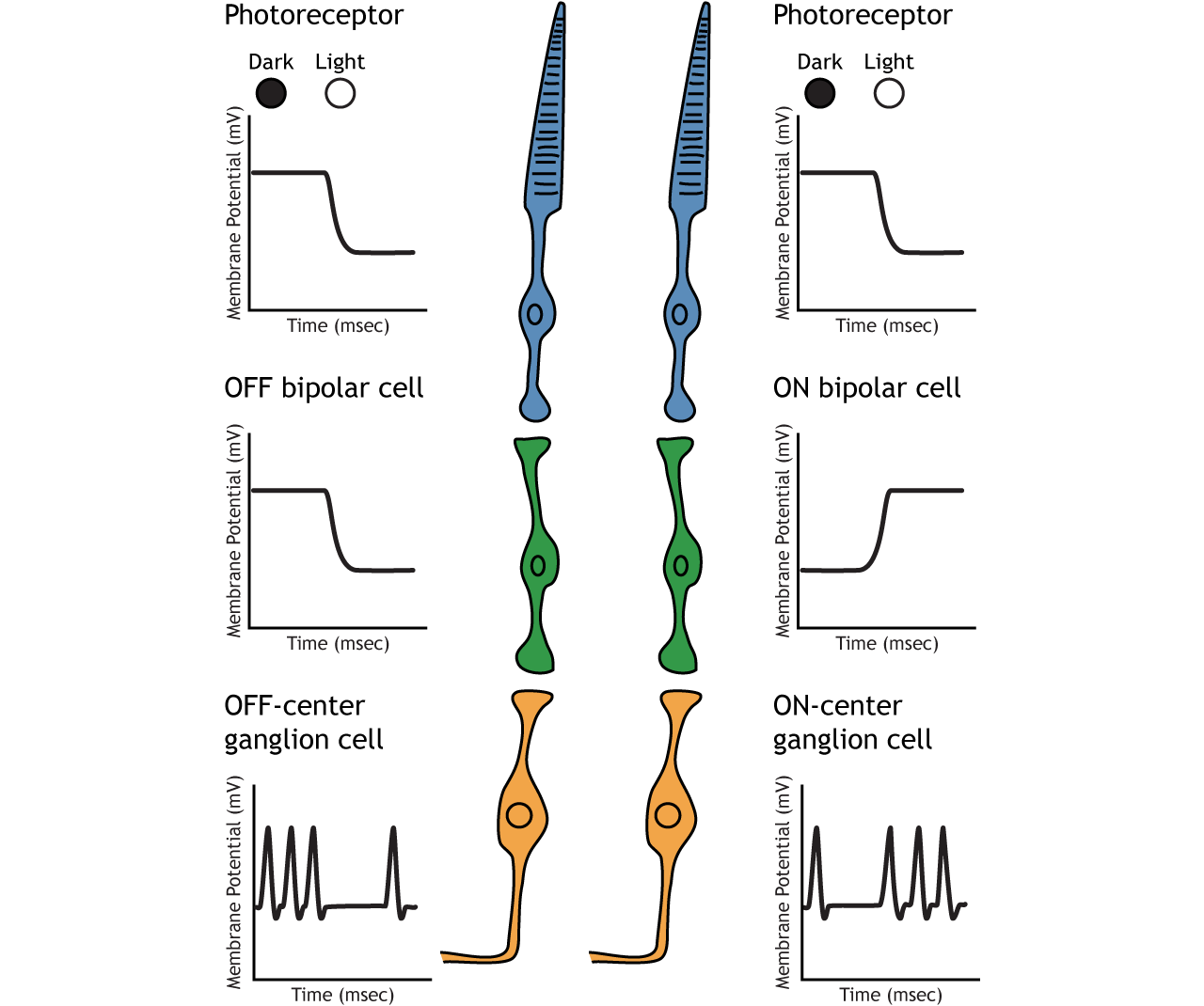
In OFF-center bipolar cells, the glutamate released by the photoreceptor is excitatory. OFF-center bipolar cells express ionotropic glutamate receptors. In the dark, the photoreceptor is depolarized, and thus releases more glutamate. The glutamate released by the photoreceptor activates the ionotropic receptors, and sodium can flow into the cell, depolarizing the membrane potential. In the light, the photoreceptor is hyperpolarized, and thus does not release glutamate. This lack of glutamate causes the ionotropic receptors to close, preventing sodium influx, hyperpolarizing the membrane potential of the OFF-center bipolar cell. One way to remember this is that OFF-center bipolar cells are excited by the dark (when the lights are OFF).
ON-center Bipolar Cells
In ON-center bipolar cells, the glutamate released by the photoreceptor is inhibitory. ON-center bipolar cells express metabotropic glutamate receptors. In the dark, the photoreceptor is depolarized, and thus releases more glutamate. The glutamate released by the photoreceptor binds to the metabotropic receptors on ON-center bipolar cells, and the G-proteins close cation channels in the membrane, stopping the influx of sodium and calcium, hyperpolarizing the membrane potential. In the light, the photoreceptor is hyperpolarized, and thus does not release glutamate. The absence of glutamate results in the ion channels being open and allowing cation influx, depolarizing the membrane potential. You can remember that ON-center bipolar cells are excited by the light (when the lights are ON).
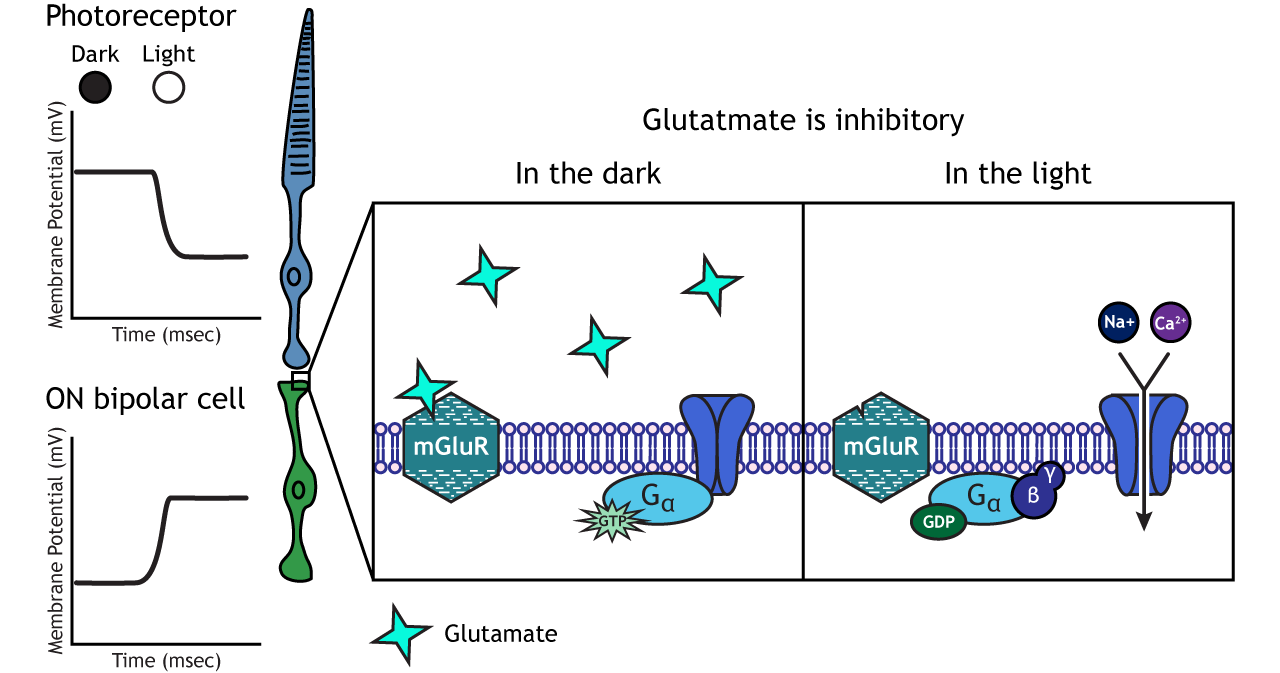
Retinal Ganglion Cells
Retinal ganglion cells are the third and last cell type that directly conveys visual sensory information, receiving inputs from the bipolar cells. OFF-center and ON-center bipolar cells synapse on OFF-center and ON-center ganglion cells, respectively. The axons of the retinal ganglion cells bundle together and form the optic nerve, which then exits the eye through the optic disk. Retinal ganglion cells are the only cell type to send information out of the retina, and they are also the only cell that fires action potentials. The ganglion cells fire in all lighting conditions, but it is the relative firing rate that encodes information about light. A move from dark to light will cause OFF-center ganglion cells to decrease their firing rate and ON-center ganglion cells to increase their firing rate.
Receptive Fields
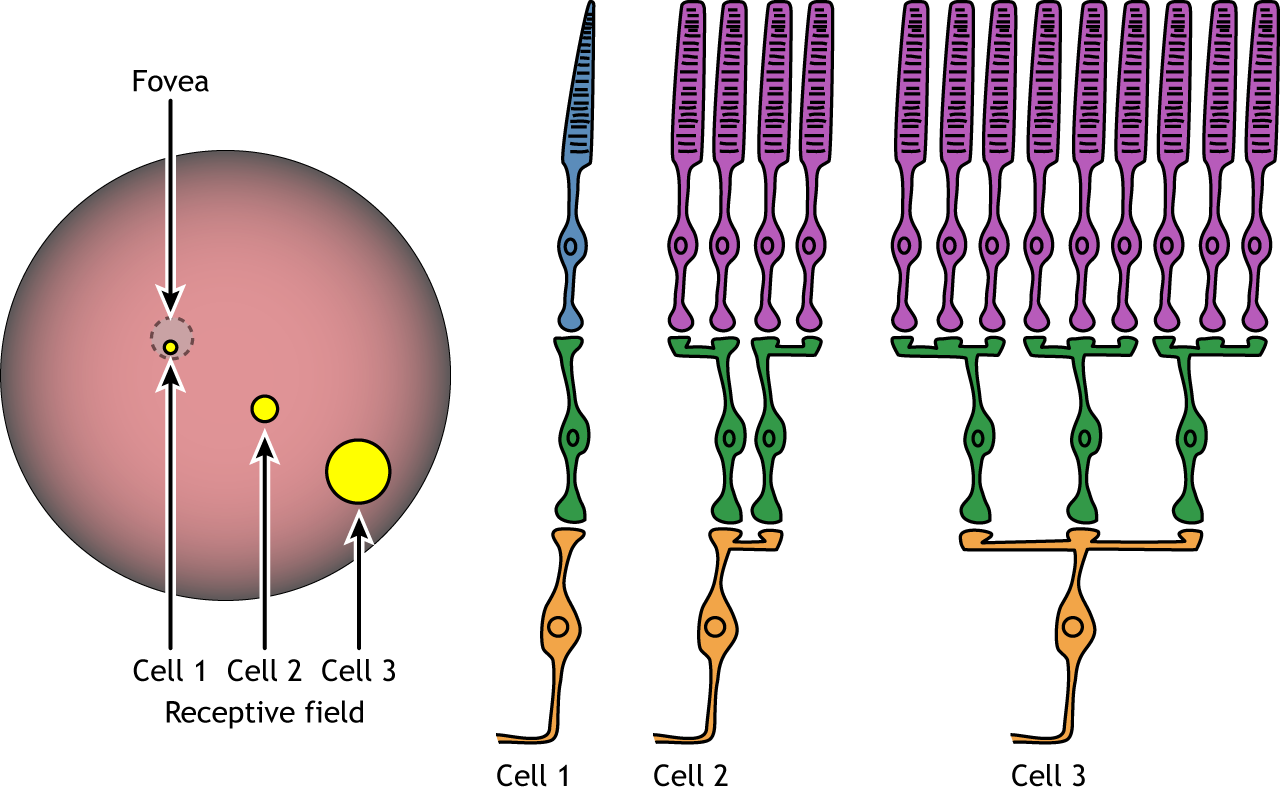
Each bipolar and ganglion cell responds to light stimulus in a specific area of the retina. This region of retina is the cell’s receptive field. Receptive fields in the retina are circular.
Size of the receptive field can vary. The fovea has smaller receptive fields than the peripheral retina. The size depends on the number of photoreceptors that synapse on a given bipolar cell and the number of bipolar cells that synapse on a given ganglion cell, also called the amount of convergence.
Receptive Field Example
Let’s use an example of an ON-center bipolar cell to look at the structure of receptive fields in the retina. The bipolar and retinal ganglion cell receptive fields are divided into two regions: the center and the surround. The center of the receptive field is a result of direct innervation between the photoreceptors, bipolar cells, and ganglion cells. If a light spot covers the center of the receptive field, the ON-center bipolar cell would depolarize, as discussed above; the light hits the photoreceptor, it hyperpolarizes, decreasing glutamate release. Less glutamate leads to less inhibition of the ON bipolar cell, and it depolarizes.
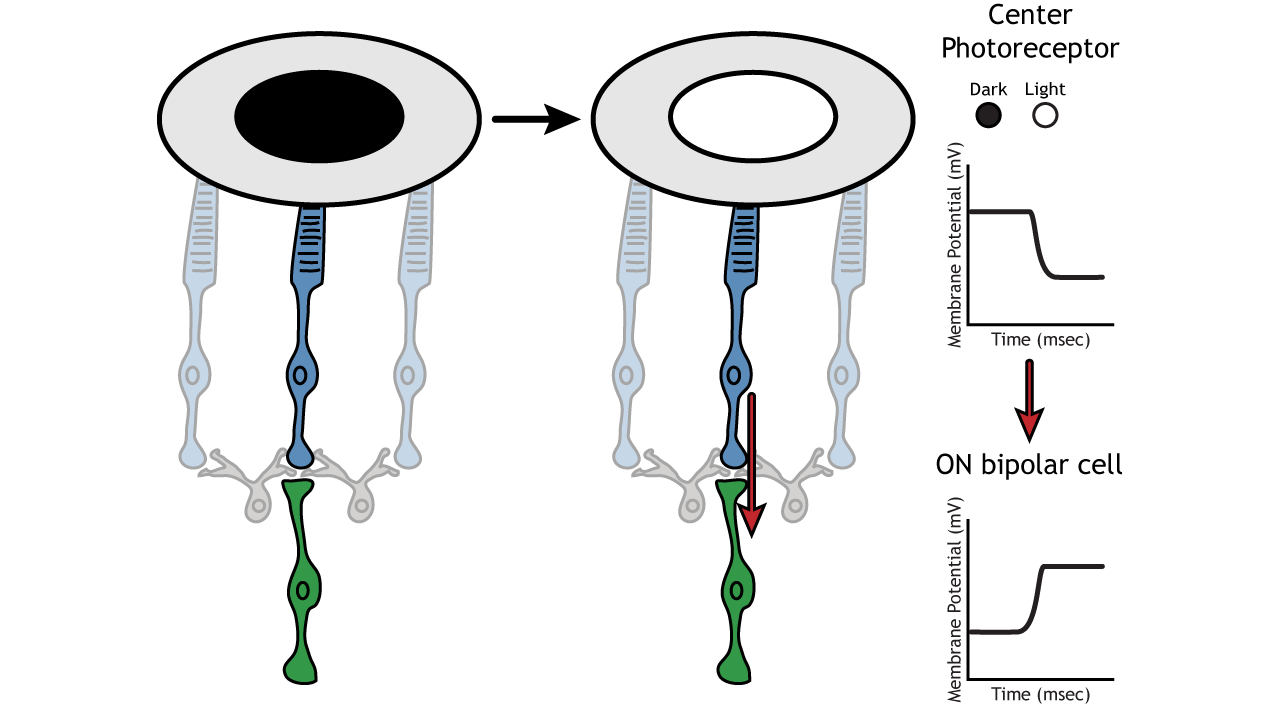
The surround portion of the receptive field is a result of indirect communication among the retinal neurons via horizontal and amacrine cells. The surround has an opposing effect on the bipolar or ganglion cell compared to the effect of the center region. That is to say, that the center and surround of the receptive field are opposite to each other. So, an ON-center bipolar or ganglion cell, can also be referred to as an “ON-center OFF-surround cell”, and an OFF-center bipolar or ganglion cell can also be referred to as an “OFF-center ON-surround cell”.
Therefore, if light covers the surround portion, the ON-center bipolar cell would respond by hyperpolarizing. The light would cause the photoreceptor in the surround to hyperpolarize. This would cause the horizontal cell to also hyperpolarize. Horizontal cells have inhibitory synaptic effects, so a hyperpolarization in the horizontal cell would lead to a depolarization in the center photoreceptor. The center photoreceptor would then cause a hyperpolarization in the ON-center bipolar cell. These effects mimic those seen when the center is in dark. So, even though the center photoreceptor is not directly experiencing a change in lighting conditions, the neurons respond as if they were moving toward dark.
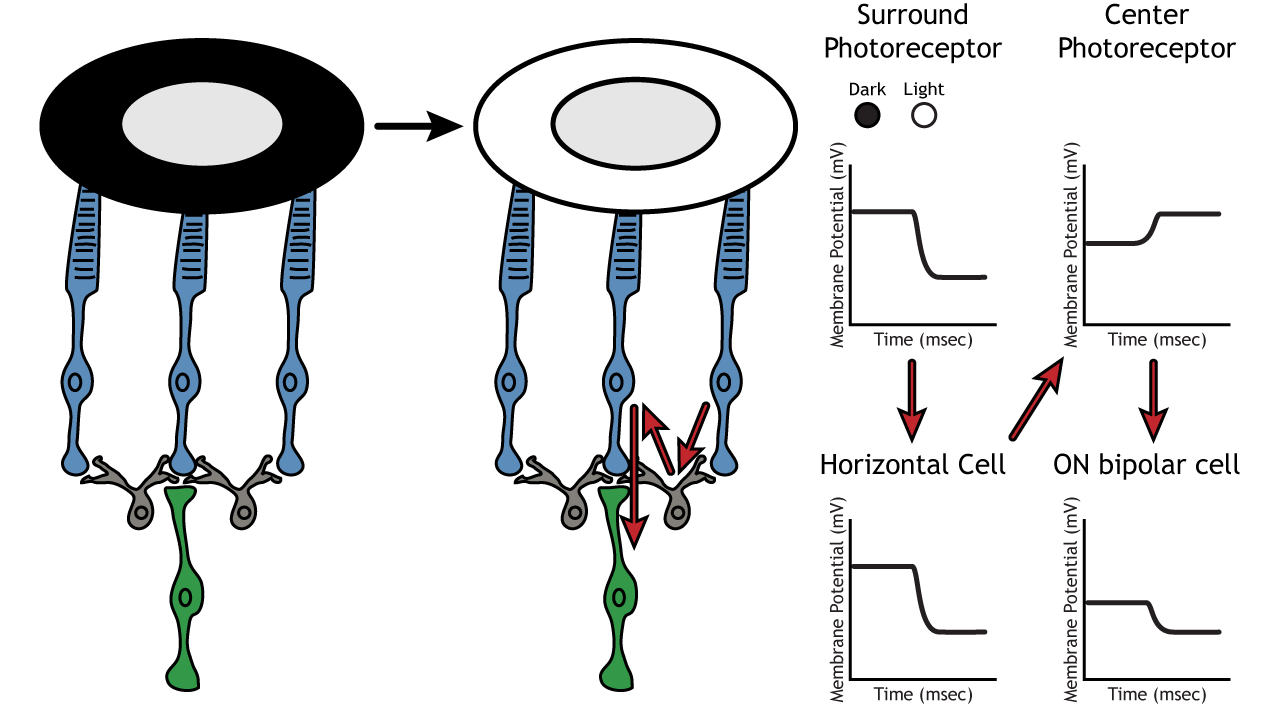
Lateral Inhibition
The center-surround structure of the receptive field is critical for lateral inhibition to occur. Lateral inhibition is the ability of the sensory systems to enhance the perception of edges of stimuli. It is important to note that the photoreceptors that are in the surround of one bipolar cell would also be in the center of a different bipolar cell. This leads to a direct synaptic effect on one bipolar cell while also having an indirect effect on another bipolar cell.
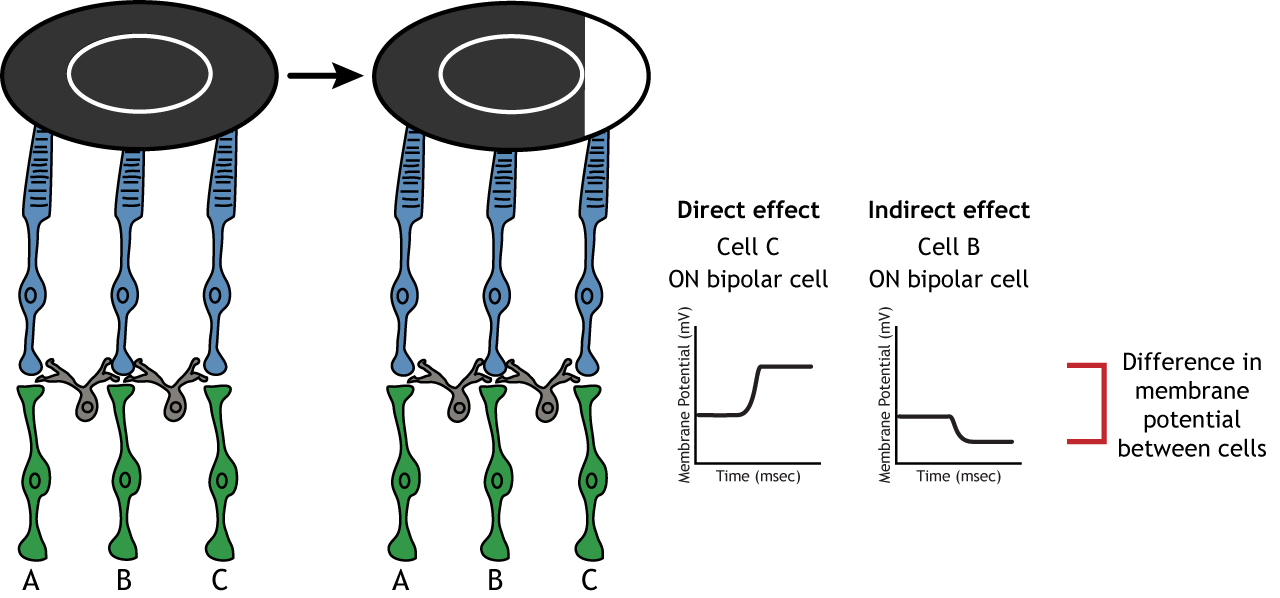
Although some of the images used here will simplify the receptive field to one cell in the center and a couple in the surround, it is important to remember that photoreceptors cover the entire surface of the retina, and the receptive field is two-dimensional. Depending on the level of convergence on the bipolar and ganglion cells, receptive fields can contain many photoreceptors.

Key Takeaways
- Photoreceptors and bipolar cells do not fire action potentials
- Photoreceptors hyperpolarize in the light
- ON bipolar cells express inhibitory metabotropic glutamate receptors
- OFF bipolar cells express excitatory ionotropic glutamate receptors
- Receptive fields are circular, have a center and a surround, and vary in size
- Receptive field structure allows for lateral inhibition to occur
Test Yourself!
Additional Review
- Compare and contrast rods and cones.
- Compare and contrast the fovea and the optic disc.
Attributions
Portions of this chapter were remixed and revised from the following sources:
- Foundations of Neuroscience by Casey Henley. The original work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License
- Open Neuroscience Initiative by Austin Lim. The original work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
Media Attributions
- EM_spectrumrevised © Philip Ronan, Gringer is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Distribution_of_Cones_and_Rods_on_Human_Retina © Jochen Burghardt is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Convergence © Casey Henley adapted by Valerie Hedges is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Fovea
Structure at the back of the eye that contains photoreceptors
Structure in the middle of the retina
Conversion of light into a neural signal and neurotransmitter release
light-sensitive components of photoreceptors
When the membrane potential gets closer to zero
When the membrane potential gets further away from zero
Postsynaptic receptors in which neurotransmitters bind and cause the opening of an ion channel. The ion channel and receptor are the same transmembrane protein.
Postsynaptic receptors in which neurotransmitters bind and cause the activation of an associated G-protein and cell signaling cascades. The affected ion channel and receptor are found on different transmembrane proteins..

{kind=link}
{kind=link}